




1. 技术全景图谱
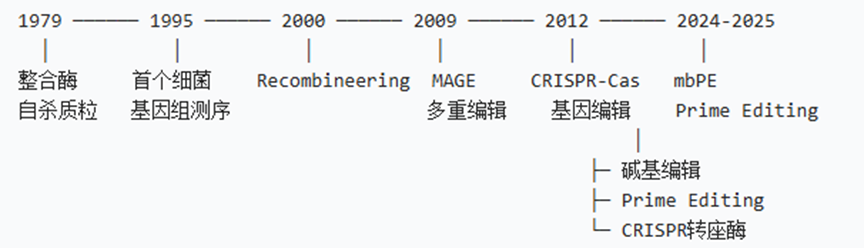
1.1 技术发展时间线
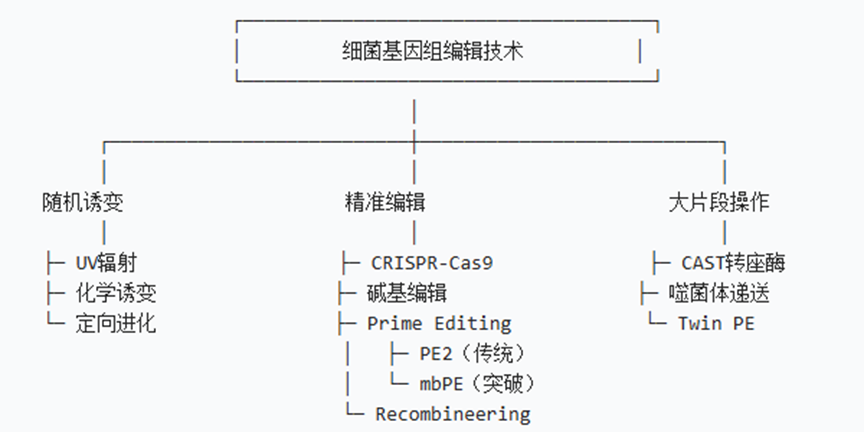
1.2 技术分类矩阵
2. 核心技术详解
2.1 CRISPR-Cas9 系统
原理:
l Cas9在sgRNA引导下在靶位点产生双链断裂(DSB)
l 细胞通过同源重组(HDR)或非同源末端连接(NHEJ)进行修复
l 在细菌中常结合“致死筛选”,未编辑细胞被清除
特点:
优势 | 局限 |
成熟稳定 | 需要供体DNA模板 |
效率高 | 多步骤操作 |
适用范围广 | SpCas9 依赖 NGG PAM |
适用菌株: 大多数细菌,包括金黄色葡萄球菌、艰难梭菌、链霉菌等
2.2 Prime Editing(引物编辑)
2.2.1 传统PE2
原理:
· Cas9切刻酶(H840A)+ 逆转录酶
· pegRNA同时提供靶向和编辑模板
· 无需外源供体DNA
在细菌中的问题:
· 选择效率低
· 需要特定遗传背景(如突变核酸外切酶),缺乏有效的正交筛选机制(如致死筛选),导致背景噪音高。
2.2.2 mbPE(Make-or-Break Prime Editing)⭐ 2025突破
核心创新:
传统PE2: Cas9切刻酶 + 需要额外筛选机制
mbPE: 野生型Cas9 + 天然致死筛选
关键参数:
· PBS长度:16 nt
· RTT长度:15 nt
· 选择效率:在肺炎链球菌中验证效率可达>90%(不同菌株中效率依赖于 DNA 修复机制与转化效率)
· 最大插入:52 bp
适用场景:
编辑类型 | 效果 |
点突变 | ✓ 高效 |
小片段删除(<100 bp) | ✓ 高效 |
小片段插入(<52 bp) | ✓ 高效 |
蛋白标签插入 | ✓ 成功 |
验证菌株: 肺炎链球菌(可扩展至其他细菌)
2.3 碱基编辑(Base Editing)
原理:
· 脱氨酶 + dCas9/Cas9切刻酶
· 直接将C→T或A→G
· 无需DSB和供体DNA
特点:
优势 | 局限 |
无DSB | 只能进行转换突变 |
效率高 | 编辑窗口限制 |
无需模板 | 主要支持转换突变(C+T,A+G)部分新型系统可实现有限的颠换,但效率和稳定性仍受限 |
2024年进展: PAM非依赖性碱基编辑,打破NGG限制
2.4 CRISPR转座酶(CAST)
原理:
· CRISPR系统指导转座酶
· RNA引导的大片段DNA插入(插入位置存在一定偏移(通常几十bp范围))
· 无需同源重组
代表系统:
系统 | 插入能力 | 来源 |
VchCAST | >10 kb | 霍乱弧菌 |
DART | kb级 | 工程化系统 |
λ-DART | kb级 | 噬菌体递送 |
2025年突破: 噬菌体λ递送DART系统,在混合菌群中实现靶向编辑
2.5 Cas12f1 迷你系统
特点:
· 蛋白大小:~500 aa(Cas9的1/3)
· 载体友好,易于包装
· 低温恢复提高效率
适用场景:
· 已报道可实现多位点编辑(通常2-3个)
· 实际能力依赖表达系统与菌株
· 载体容量受限的情况
3. 技术选型决策
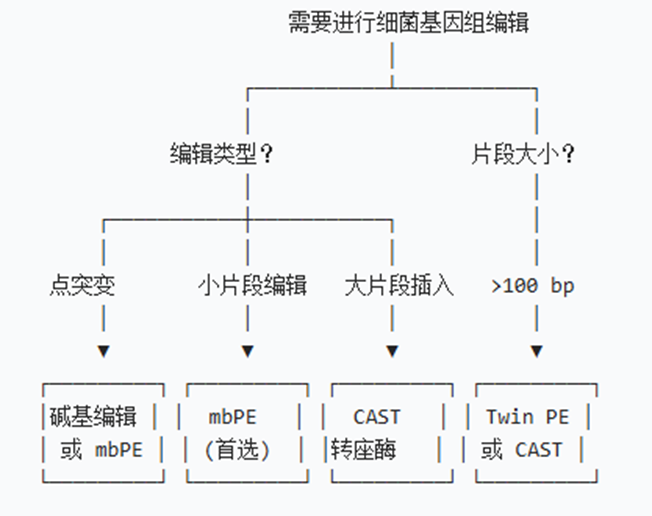
场景1:单碱基突变
√如果仅需C+T或A+G
推荐:碱基编辑(简单高效)
√如果需要任意碱基替换
推荐:mbPE
场景2:小片段编辑(<52 bp)
插入 or 删除?
├─ 删除 → mbPE(<52 bp)
└─ 插入 → mbPE(<52 bp)或 CAST(>52 bp)
场景3:大片段操作(>1 kb)
插入 or 删除 or 替换?
├─ 插入 → CAST转座酶 或 λ-DART
├─ 删除 → CRISPR-Cas9 + 同源重组
└─ 替换 → Twin PE 或 CAST
场景4:多重编辑(>2个位点)
位点数量?
├─ 2-3个 → Cas12f1 或 mbPE库
└─ >3个 → MAGE 或 高通量CRISPR
场景5:困难菌株
菌株类型?
├─ 转化困难 → SSAPs库(宽宿主范围)
├─ RecA缺陷 → mbPE(RecA非依赖)
└─ 混合菌群 → λ-DART(噬菌体递送)
4. 编辑效率对比
4.1 各技术效率汇总
技术 | 点突变 | 删除 | 插入 | 多重编辑 |
CRISPR-Cas9 | 70-95% | 80-99% | 50-90% | 中等(依赖RecT/R) |
碱基编辑 | 90-99% | ✗ | ✗ | ✗ |
mbPE | >93% | >93% | >93% | 高通量 |
CAST | 低 | 低 | 50-99% | 支持 |
SSAPs+Cas9 | >70% | 高 | 中等 | 支持 |
Cas12f1 | 高 | 高 | 中等 | 3位点 |
5. 应用场景匹配
5.1 按编辑目的分类
功能研究
目的 | 推荐技术 | 理由 | |
基因敲除 | CRISPR-Cas9 | 成熟可靠 | |
点突变验证 | mbPE | 高精度,无痕 | |
功能域删除 | mbPE | 精确边界 | |
蛋白标记 | mbPE | 标签插入 | |
菌株改造
目的 | 推荐技术 | 理由 |
代谢通路优化 | 多重编辑 | 同时修改多基因 |
抗性基因插入 | CAST | 大片段稳定整合 |
启动子替换 | mbPE/CAST | 精确替换 |
益生菌改造 | mbPE | 无痕编辑 |
高通量筛选
目的 | 推荐技术 | 理由 |
全基因组功能筛选 | CRISPRi-seq | 覆盖广 |
突变体库构建 | mbPE库 | 高通量 |
酶定向进化 | 碱基编辑/mbPE | 多样性 |
5.2 按菌株类型分类
模式菌株
菌株 | 推荐技术 | 备注 |
大肠杆菌 | 全部可用 | 工具最完善 |
枯草芽孢杆菌 | MAD7/mbPE | CRISPR-MAD7成熟 |
肺炎链球菌 | mbPE | 针对性优化 |
金黄色葡萄球菌 | CRISPR-Cas9/SSAPs | 已验证 |
工业菌株
菌株 | 推荐技术 | 挑战 |
谷氨酸棒杆菌 | FnCpf1/mbPE | SpCas9有毒 |
链霉菌 | CRISPR-Cas9 | 转化效率低 |
梭菌 | CRISPR-Cas9 | 厌氧条件 |
益生菌
菌株 | 推荐技术 | 应用 |
乳酸菌 | CRISPR-Cas9 | 发酵优化 |
双歧杆菌 | SSAPs | 转化困难 |
Akkermansia | 待开发 | 研究热点 |
6.技术风险与限制
CRISPR-Cas9:可能产生脱靶或细胞毒性
mbPE:依赖特定菌株背景
CAST:插入位点存在偏移
Base Editing:存在编辑窗口限制
总结
技术 | 是否DSB | 是否模板 | 可编辑类型 | 插入能力 | 适合场景 |
CRISPR-Cas9 | √ | √ | 全部 | Kb级 | 通用编辑 |
碱基编辑 | × | × | 点突变 | × | 快速突变 |
mbPE | × | × | 小片段 | <52bp | 精准编辑 |
CAST | × | × | 插入 | >10kb | 大片段工程 |
附录
A. 关键文献索引
技术 | 关键文献 | 年份 |
CRISPR-Cas9细菌编辑 | Jiang et al., Nat Biotechnol | 2013 |
Prime Editing | Anzalone et al., Nature | 2019 |
mbPE | Rengifo-Gonzalez et al., Nat Commun | 2025 |
SSAPs库 | Filsinger et al., PNAS | 2025 |
λ-DART | Roberts et al., PNAS | 2025 |
Cas12f1多重编辑 | Lim et al., J Microbiol Biotechnol | 2024 |
B. 载体资源
载体类型 | 来源 | 用途 |
mbPE pegRNA载体 | pVL4132/4133/4393 | 肺炎链球菌编辑 |
CAST载体 | Addgene | 大片段插入 |
pORTMAGE系列 | Addgene | SSAPs重组工程 |
C. 设计工具
工具 | 用途 | 链接 |
PrimeDesign | pegRNA设计 | 在线工具 |
CHOPCHOP | sgRNA设计 | 在线工具 |
CRISPRi-Seq | 全基因组筛选 | 分析流程 |
参考文献:
[1] 李琦, 武美贤, 郭清华, 邵悠然, 杨俊杰, 蒋宇, 杨晟. 细菌基因组编辑技术进展[J]. 生命科学, 2019, 31(5): 473-492. DOI: 10.13376/j.cbls/2019201.
[2] Rengifo-Gonzalez M, Mazzuoli MV, Janssen AB, Rueff AS, Burnier J, Liu X, Veening JW. Make-or-break prime editing for genome engineering in Streptococcus pneumoniae[J]. Nature Communications, 2025, 16: 3796. DOI: 10.1038/s41467-025-59068-8.




